In Vitro Hippocampal Electrophysiology and in Vivo Quantitative EEG Revealed Robust Neurophysiological Effects of the Antivertigo-Agent Vertigoheel® in a Rat Study
Wilfried Dimpfel1*, Bernd Seilheimer2, Leonie Schombert31Justus Liebig University Giessen, Giessen, Germany. 2Biologische Heilmittel Heel GmbH, Baden-Baden, Germany. 3NeuroCode AG, Wetzlar, Germany.
Abstract
Vertigo is a common symptom with impact on daily life. Vertigoheel® (VH-04) has demonstrated to be effective for Vertigo in former studies. This paper aims to investigate the mode of action of the medicinal product VH-04 in the rat brain. In an in vitro study neurophysiological recording from hippocampal slices from adult male Sprague Dawley® rats was performed in order to substantiate a possible direct effect on the brain of VH-04 in different concentrations. In an in vivo cross-over study with 11 Fischer 344® rats, a neurophysiological method was applied to systemically analyse VH-04’s activity in the rat brain. This method combines quantitative assessments of telemetrically transmitted field potentials after drug treatment with subsequent discriminant analysis to classify the compound. The database used for the analysis of classification contained numerous chemicals and medicinal products of different dosages, all tested in the same paradigm, which is continuous wireless monitoring of the EEG of freely moving rats before and after drug intake. Following single stimuli on the Schaffer collaterals in the presence of VH-04 in different concentrations, in vitro responses of pyramidal cells increased depending on the VH-04 concentration (0.25 - 4 ml/L). Results were statistically significant for concentrations above 2.5 ml/L. Long-term potentiation was only marginally affected. Out of several specific glutamate receptor antagonists the effect of VH-04 was only antagonized by AMPA and kainic acid receptor-mediated signalling. Their enhancement indicates better information processing in the hippocampus, a brain structure primarily involved in memory processes. The in vivo characterisation of VH-04-induced changes in EEG-signatures of four brain areas (the frontal cortex (FC), the hippocampus (HC), the striatum (ST) and the reticular formation (RF)) revealed a dose-dependent attenuation of delta, theta, alpha 2 and beta 1 waves. The subsequent discriminant function analysis classified the VH-04 EEG-signature into a subset of cognition-enhancing medicinal products.
Keywords
EEG, Rat Model, Central Nervous System, Vertigo, VH-04, Neurophysiology, Hippocampus, Multitarget, Fast Fourier Transformation
Share and Cite:
Dimpfel, W. , Seilheimer, B. and Schombert, L. (2019) In Vitro Hippocampal Electrophysiology and in Vivo Quantitative EEG Revealed Robust Neurophysiological Effects of the Antivertigo-Agent Vertigoheel® in a Rat Study. Neuroscience and Medicine, 10, 407-425. doi: 10.4236/nm.2019.104030.
- Introduction
The terms vertigo, dizziness, and unsteadiness are used inconsistently by the public as well as professionals and there is a general demand for a more precise definition of vertigo in clinical practice as shown by Blakley et al. [1]. Anyhow, the lifetime prevalence for vertigo of 7.8% in the general population was estimated with a significant rise with age as published by Neuhauser et al. [2]. Vertigo is a symptom, and depending on its origin, vertigo is distinguished between vestibular and non-vestibular vertigo. Vestibular vertigo is distinguished between peripheral and central disorders. To the peripheral vestibular disorders belong vestibular neuritis, benign paroxysmal positional vertigo, Ménière’s disease and bilateral vestibulopathy. Non-vestibular vertigo has different etiologies such as metabolic disorders (diabetes mellitus), drug-induced dizziness, orthostatic dysregulation, stroke affecting other cerebral areas and psychiatric disorders (anxiety, depression). Although a number of specific conditions are defined that can cause vertigo, the condition is often multifactorial mainly in, but not restricted to the elderly [2] complicating the diagnosis and treatment of the symptom. Therefore, the diagnostic and therapeutic approach must be multi-systemic and oriented to the visual, proprioceptive, and vestibular systems according to Fernández et al. [3].
Several studies have investigated the medicinal product VH-04. In a randomized double-blind controlled clinical study Weiser et al. [4] compare the treatment of vertigo to the treatment of betahistine. It could be demonstrated on 105 patients in 15 study centers, receiving either VH-04 or betahistine hydrochloride, that both treatments reduced the frequency, duration, and intensity of vertigo attacks during a six-week treatment period. Klopp et al. [5] conducted a non-randomised open label study with 32 patients, revealing that parameters of subcutaneous microcirculation as evidenced by arteriolar and venular erythrocyte flow rates, were increased via VH-04. In vitro studies demonstrated a dose-dependent vasorelaxation in isolated pre-contracted carotid artery rings by phenylepinephrine. Data on vasorelaxation and microcirculatory improvements may indicate joint actions providing a first hint towards an antivertiginous effect of VH-04, but do not explain the combinatorial effects its multicomponent ingredients. Unraveling VH-04’s mode of action remains a challenge because it possesses a complex repertoire of chemicals that belong to a variety of substance classes, which is in contrast to defined single drug combinations. Therefore, activity monitoring requires a reconsideration of the conventional methods. According to Wink [6], the pharmacological activity of multicomponent mixtures cannot be assigned to a single substance, and the contained phytochemicals typically act in an unspecific and widespread manner. It is the summation of activities that possibly leads to a potentiation of effects and promotes a prominent outcome. Conversely, the fractionation or isolation of principal constituents from extracts has been shown to end up in a loss of previously detected activities according to Keith et al. [7]. Numerous theories propose that the interplay and interference of single components in a mixture is the rationale for the advantageous effects of multicomponent medicinal products. The basic concepts of joint actions and interactions can be summarised as three core processes: addition, synergism and antagonism [8] [9] [10] [11] [12]. However, the response to substances in a highly interconnected biological system, such as the brain, is very complex and rarely linear. There are various factors that can influence the therapeutic efficacy of a substance, such as the effective intracellular concentration of compounds at the target cell or organ (e.g. bioavailability, bioconversion, pharmacokinetics), the chemical and physical microenvironment (e.g. polarity) at the interaction site, the type of molecular target (e.g. single or multiple molecules, cellular structures), and the general health of the target cells. Holistic activity monitoring of complex multicomponent medicinal products requires an implementation of new strategies that allow for an integrated overview of participating molecular processes.
- Material and Methods
2.1. Material
VH-04 (Vertigoheel®) is a multicomponent, multitarget medicinal product that has been established in the treatment of vertigo and dizziness of various origins for many years in numerous countries around the world. For injection (1.1 ml) it is a clear colourless, odourless solution with multiple components (Table 1). For human use it is injectable either intramuscular (i.m.), subcutaneous (s.c.), intracutaneous (i.c.) or intravenous (i.v.). In this study 0.5% ethanol v/v was added to

Table 1. Components, amount and dilution of VH-04.
the study medicinal product. Dosages of 0.5 ml, 1.0 ml or 2.0 ml per kg body weight (BW) of VH-04 were injected intraperitoneally (i.p.). The vehicle control received 0.9% natrium chloride (NaCl) in equal volumes using a separate vehicle control group for each dose level with n = 11 per group.
An overview of glutamatergic receptor-specific chemicals is given in Table 2. All chemicals were bought from BioTrend Chemicals AG, Wangen/Zürich, Switzerland.
2.2. Animals and Trial Setting
In vivo studies were performed on twenty-two eight-month-old Fisher 344® rats (Charles River Laboratories, Sulzfeld, Germany) which were kept in sterile filter-top cages on a reversed day/night cycle and provided with food and water Ad libidum. The implantation of bipolar concentric steel electrodes into the FC, the HC, the ST and the RF were achieved by screws drilled into the scull and fixated by red dental cement. Signals were wirelessly transmitted by a radio-telemetric system (TSE-Systems GmbH, Bad Homburg, Germany), using 40 megahertz (MHz) as a carrier frequency, and were amplified and processed to give the desired power spectra of a 0.25 Hertz (Hz) resolution according to Dimpfel et al. [13]. The animals were granted a two-week recovery period before drug treatment. The reversed day and night cycle was continued for the whole study. The study was performed during the four weeks after surgery. A time course is documented in Figure 1. After a forty-five-minute pre-drug baseline recording, single doses of VH-04 or vehicle control were administered as indicated (Figure 2). After a five-minute recovery interval, EEG-signals were then recorded for one hour. Different doses of VH-04 were tested in a crossover design. A total of eleven animals were exposed to one dose of the medicinal product per experiment followed by a one-week drug-free interval. One animal from the highest dose group was excluded due to technical issues. Principles of laboratory (NeuroCode

Table 2. Overview of glutamatergic receptor antagonists, their chemical names, their batch numbers and their synonyms.
Figure 1. Time course of experimental series.
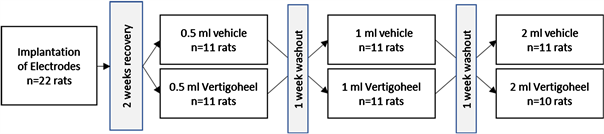
Figure 2. Details of administration.

AG, Wetzlar, Germany) animal care were followed in all trials. All animal procedures were performed according to German Animal Health Guidelines. Animal experiments were approved by the government authority Regierungspräsidium Giessen dated 25th Febr. 2010.
2.3. Procedures
2.3.1. Hippocampal Slices in Vitro
HC slices were obtained from forty-two adult male Sprague Dawley® rats (Charles River Laboratories, Sulzfeld, Germany). The rats were kept under a reversed day/night cycle for two weeks prior to the experiments, in order to record in vitro activity in slices during the active phase of their circadian rhythm according to Dimpfel et al. [14] and Dimpfel [15]. Animals were sacrificed, brains removed, and hippocampi isolated under micro-stereoscopic view. The midsection of the HC is fixed to the table of a vibrating microtome using a cyanoacrylate adhesive, submerged in chilled bicarbonate-buffered saline, and cut into slices of 400 μ thickness. All slices are pre-incubated for at least one hour in Carbogen saturated artificial cerebrospinal fluid (ACSF; 124 mM NaCl, 5 mM KCl, 2 mM CaCl2, 2 mM MgSO4, 26 mM NaHCO3, 10 mM glucose; pH = 7.4) in a pre-chamber before use, according to Dimpfel [15]. The methodology has been described earlier by Dimpfel [16] in more detail. During the experiment, slices were held and treated in a special super-fusion chamber (List Electronics, Darmstadt, Germany) according to Haas et al. [17] and kept at 35˚C as reported in the literature [18]. Four slices were used from one rat per day under one of the test conditions, either control or different concentrations of the test preparation. After obtaining stable responses to singular stimulus (SS), long-term potentiation was induced by applying a theta burst stimulus (TBS) type pattern [19].
2.3.2. EEG Recording in Vivo
Animals were day-night converted to allow recording during the active phase. EEG-recordings were performed at the same time of day to exclude circadian effects. During recording, rats were able to move freely but had no access to food in order to avoid artefacts caused by chewing. Signals were transmitted and processed to give power spectra of a 0.25 Hz resolution as described earlier in Dimpfel [20]. Briefly, artefacts were automatically rejected and signals of 4 s units were collected and Fast Fourier transformed using a Hanning window, 512 Hz sampling frequency. Four values were averaged to give a final sampling frequency of 128 Hz. The resulting electrical power spectra were divided into six defined frequency ranges (Table 3). Spectra were averaged in three-minute intervals during recording in order to supervise the data acquisition and in sixty-minute intervals during offline analysis for final evaluation.
2.4. Discriminant Analysis
The effects of VH-04 were further compared with those of reference drugs using twenty-four variables, six frequencies times four brain areas, by linear discriminant analysis according to Fischer as described previously in Dimpfel [21]. The results of the first three discriminant functions were projected into space, on X-, Y- and Z-axis, whereas the results from the next three discriminant functions are documented by means of an additive mixture of the colours red, green and blue (RGB). Previous work [20] [22] [23] revealed that synthetic medicinal products interacting with the special neurotransmitters (e.g. norepinephrine, dopamine, acetylcholine, etc.) show particular patterns of frequency changes. In addition, these frequency changes allowed an assignment to different indication classes (e.g. anxiolytics, anti-depressants, analgesics, etc.). The different actions of medicinal products were analysed by evaluating all frequency changes in the four brain regions at the same time in a statistically significant manner. The results were represented in six dimensions. Projecting the VH-04 data into this six-dimensional graph allowed the positioning of the study medicinal product within a selection of twelve indication-specific synthetic reference drugs (Table 4). This positioning is useful because it quantitatively describes the relational similarities and differences to all plotted medicinal products.
2.5. Statistical Analysis
Averages of EEG-signals in µV2 were expressed as percent of pre-drug baseline values, and data of all animals per group were summarised as mean ± standard

Table 3. Brain wave names, their frequency ranges and colour coding.

Table 4. Listing of reference compounds used for discriminant analysis tested previously by the experimenter. Doses of drugs are given. Compounds are administered i.p. All drugs are analysed for 60 minutes after i.p. administration except for Phenytoin (65 to 125 minutes).
error of the mean (SEM) during the first hour after administration. For the in vitro and in vivo experiments, significant differences between experimental medicinal product and vehicle control were identified by Wilcoxon Mann-Whitney U-test with no adjustments for multiple comparison. p-values of less than 0.05 were considered significant.
- Results
3.1. Hippocampal Slices in Vitro
Following electric single shock stimulation (SS) to the Schaffer collaterals, responses of the pyramidal cells increased concentration dependently in the presence of VH-04 (Figure 3). The population spike amplitude increased, reaching its peak at 2 ml/L of VH-04, amplitude increases from 1.07 to 1.73 mV. There are significant statistical differences between this increase rate and the control value without medicinal product (p < 0.01). The next higher dosage of 4 ml/L of VH-04 showed somewhat lower values and thus a trend towards a “bell-shaped” curve. Following electric theta burst stimulation (TBS) the presence of VH-04 only led to a weak trend in increases of the population spike amplitude, from 2.06 mV to 2.24 mV.
In order to investigate particular targets of VH-04, several glutamate receptor antagonists were added to the superfusion medium of the HC slice (Figure 4). Appropriate dosages were determined via pilot studies. Effective concentrations of VH-04 were taken from the first series of experiments depicted in Figure 3, with 1 ml/L for VH-04. In order to test a possible interference of VH-04 with NMDA receptor activated signal changes, the glutamatergic neurotransmission
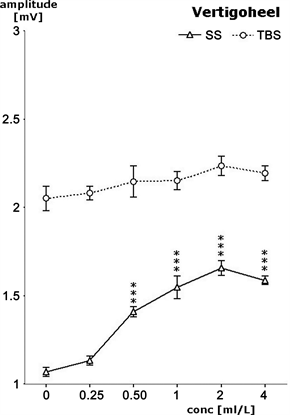
Figure 3. Concentration dependent effects of Vertigoheel (VH-04) on pyramidal cell activity in terms of changes of population spike amplitudes after dilution of the commercial preparation as ml per liter (X-axis). Results from single slices as obtained after single stimuli (SS) or after theta burst stimuli (TBS). Data are given as mean ± S.E.M. of n = 6 slices. Amplitude of the signal is given as voltage in mV (Y-axis). Lower curve represents changes after single shock stimulation, upper curve represents changes after theta burst stimulation. Statistical significance between the effects of control and VH-04 is marked as follows: ***p < 0.01 according to Wilcoxon, Mann and Whitney.
was modulated by CGS 19755, a very potent and selective NMDA receptor antagonist according to Bennett et al. [24]. In the presence of 250 nM CGS 19755 alone, only minor decreases of the signal amplitude could be measured (Figure 4). A concentration of 250 nM did not antagonize the amplitude increase in the presence of VH-04 after a SS (Figure 5). Thus, no interference with the NMDA receptor-mediated signal transmission could be observed in the presence of VH-04. In order to test a possible interference of VH-04 with AMPA receptor-activated signal changes, glutamatergic neurotransmission was modulated by NBQX, a very potent and selective competitive AMPA receptor antagonist according to Gill et al. [25]. In the presence of 50 nM NBQX alone, no change in the signal amplitude could be measured. However, a concentration of 50 nM blocked the significant enhancement of the population spike amplitude in the presence of VH-04 after SS (Figure 5). Thus, for VH-04 a clear interference with the AMPA receptor-mediated signal transmission was observed. To test a possible interference of both medicinal products with non-competitive AMPA glutamate receptor activated signal changes, glutamatergic neurotransmission was modulated by GYKI 52466, a very potent and selective non-competitive AMPA receptor antagonist according to Szabados et al. [26]. In the presence of 500 nM GYKI 52466 alone, only very minor decreases of the signal amplitude were measured. A concentration of 500 nM of GYKI 52466 did not antagonize the increased amplitude in the presence of VH-04 after SS (Figure 5). Thus, no antagonism with the non-competitive AMPA receptor mediated signal transmission was observed for VH-04 after SS. In the presence of TBS, however, a slight, statistically insignificant increase of the population spike amplitude was seen (Figure 5). Nearly identical results were obtained in the presence of SYM 2206 according to Pelletier et al. [27], another non-competitive AMPA receptor antagonist (Figure 5). In order to test a possible interference of VH-04 with kainate receptors—glutamatergic neurotransmission was modulated by UBP 30, a very potent and selective competitive kainate receptor antagonist according to More et al. [28]. In the presence of 50 nM UBP 301 alone, no change in the signal amplitude could be measured (Figure 4). Nevertheless, a concentration of 50 nM antagonized completely the statistically significant enhancement of the population spike amplitude in the presence of VH-04 after SS (Figure 5). Thus, for VH-04 a clear interference with the kainate acid receptor mediated signal transmission was observed. To determine a possible interference of VH-04 with a class I metabotropic glutamate receptor glutamatergic neurotransmission was modulated by YM 298198, a very potent and selective metabotropic glutamate receptor I antagonist [29]. During superfusion with 50 nM YM 298198 alone, only a minor decrease of the signal amplitude after TBS could be measured (Figure 4). Amplitudes following SS were
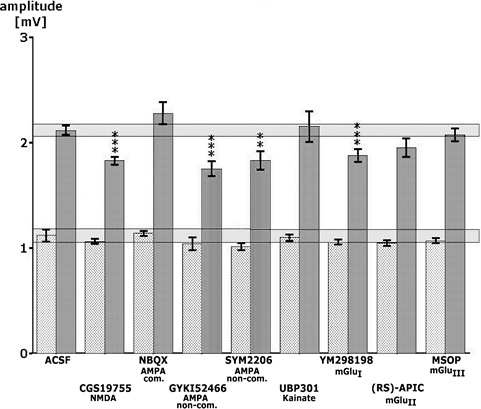
Figure 4. The effects of low concentrations of several glutamate receptor antagonists acting on NMDA receptor, AMPA receptor, Kainate receptor and 3 metabotropic glutamate receptors (Glu I, II, III) in comparison to the effects of artificial cerebrospinal fluid (ACSF). Data for SS are represented by the lightly shadowed bars and data for TBS are represented by dark-grey bars. The range of control data is shaded horizontally. Statistical significance between the effects of control and various receptor antagonists is marked as follows: **p < 0.05, ***p < 0.01 according to Wilcoxon, Mann and Whitney.
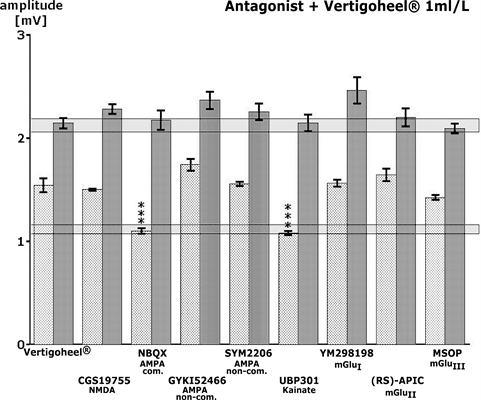
Figure 5. The effects of Vertigoheel VH-04 (1 ml/L) in the presence of 8 glutamate receptor antagonists. Data for SS are represented by the lightly shaded bars, data for TBS are represented by the dark-grey bars. The range of control data is shadowed horizontally. Statistical significance between the effects of VH-04 alone and in the presence of the receptor active compound is marked: ***p < 0.01 according to Wilcoxon, Mann and Whitney.
not changed in the presence of VH-04 plus the antagonist. However, during TBS a small increase of the amplitude was seen in the presence of VH-04 plus the antagonist. A class II metabotropic glutamate receptor antagonist (RS)-APICA [30] at a concentration of 100 nM alone did not inhibit the signal amplitude (Figure 4). The effect of VH-04 after SS was not influenced (Figure 5). A class III metabotropic glutamate receptor antagonist MSOP [31] at a concentration of 50 nM alone did not change the signal amplitude after SS or TBS. The effect of VH-04 after SS was not changed by this compound (Figure 5). In summary, the effect of VH-04 was only antagonized by a competitive AMPA and a kainate receptor antagonist.
3.2. EEG Recordings in Vivo
The changes of electrical activity at several locations of the brain recorded in the presence of VH-04 in different dosages (the so-called electropharmacogram, EPG) were recorded continuously for one hour after drug administration following the forty-five minutes lasting baseline. Changes in spectral power within the four brain regions following physiological saline administration could hardly be detected, which is documented for the first hour after administration in Figure 6. Spectral frequency changes in the presence of the lowest dosage of VH-04 (0.5 ml/kg i.p.) were dominantly seen in the alpha 2 frequency in all brain areas, but did not reach statistical significance in comparison to saline injection (Figure 6). Doubling of the dosage (1.0 ml/kg BW) provided a more profound picture of spectral frequency changes. Alpha 2 frequencies were attenuated in all brain regions in a statistically significant manner. The strongest changes, however, were observed in the FC and HC. Here, all frequencies were attenuated in a significant manner except for alpha 1 and beta 2. Statistically significant attenuation of alpha 2 and beta 1 waves were also observed in the ST. An administration of the highest dosage of 2.0 ml/kg led to similar spectral frequency changes as observed for the lower dosage of 1 ml/kg. Highly significant attenuation of electrical power was seen in the FC, the HC and the ST with respect to delta, theta, alpha 2 and beta 1 frequencies during the first hour after administration. A statistically significant attenuation of alpha 1 waves occurred in the HC and the RF only with the highest dosage (Figure 6).
Similarities between the 1.0 ml/kg frequency change pattern exhibited by VH-04 and those of reference compounds, which had been tested earlier under identical conditions, were observed. Two medicinal products, Metanicotine, a stable nicotine-analogon acting at the cholinergic system, and Tacrine, an inhibitor of acetylcholinesterase, turned out to produce very similar changes in the frequency content of field potentials. Furthermore, caffeine also induced a comparable frequency pattern, shown in Figure 7.
A comparison between the complicated patterns of frequency changes was performed using discriminant analysis. Twenty-four variables, and the patterns of 12 clinically proven reference medicinal products, which had been tested earlier under similar conditions, were obtained from the data-base. The projection of the calculation results led to an obvious separation of the compounds’ action on
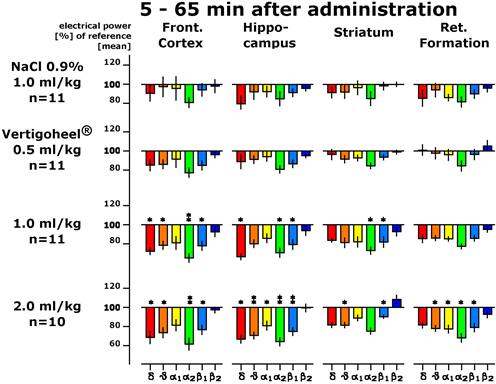
Figure 6. EPG showing the dose-dependent effect of Vertigoheel (VH-04) with respect to four brain areas and six frequency ranges from delta (left) through theta, alpha 1, alpha 2, beta 1 to beta 2. The ordinate shows electrical power changes in percent of baseline values. Bars pointing downwards depict the attenuation of power. Asterisks depict following statistical significance: *p < 0.05; **p < 0.01 according to Wilcoxon, Mann and Whitney.
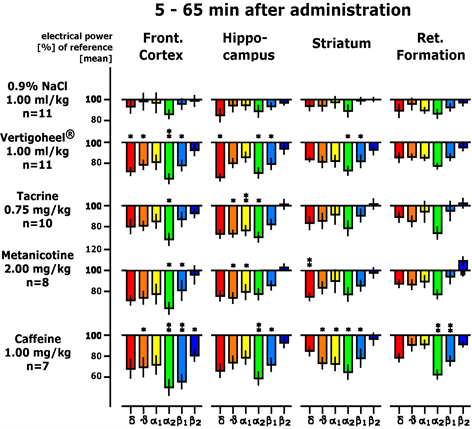
Figure 7. Comparison of the effects of Vertigoheel (VH-04) to frequency patterns evidenced by reference medicinal products in earlier trials using the identical methodology. Please note very similar frequency pattern of VH-04 to reference medicinal products. Common effects are seen with respect to alpha 2 and delta waves (labelled green and red). Asterisks depict following statistical significance: *p < 0.05; **p < 0.01 according to Wilcoxon, Mann and Whitney.
the brain based on changes in the frequency content of the EPG. The three spatial coordinates were used for the first three discriminant functions and the results of the second three discriminant functions were depicted in the colours red, green and blue (so-called RGB mode). Thus, indication-specific activity changes can be visualised in a more interpretable manner by grouping the projection of reference compounds. Figure 8 documents this indication-specific grouping. These groupings correspond quite well to pharmacological textbook indications. Firstly, three-dimensional space evidences similarity or dissimilarity of drug action. Additionally, equal or similar colours suggest similar mechanisms of action. The position and colour of reference compounds with known clinical indications are listed in Table 4. Medicinal products injected i.p. are analysed for their action during the next hour after administration. The preparation VH-04 is analysed during the time of 5 - 65 minutes after administration. VH-04 is represented by one dosage of 1.0 ml/kg and marked as “Vertigoheel” (VH-04). The EPG of VH-04 is projected in close vicinity to the cognition-enhancing medicinal products Tacrine and Metanicotine. VH-04 is also projected near caffeine, as stimulating chemical.
Concomitantly with the telemetric recording, motion of the freely moving rats was monitored. There were no obvious changes in motion in the presence of VH-04 compared to control of 0.9% NaCl. Table 5 depicts changes in motion

Figure 8. Discriminant analysis of EPG providing similar patterns of spectral frequency changes for similar clinical indications. Compounds having a similar effect appear in close vicinity to each other (see X-, Y- and Z-axis). Great similarity with respect to space and colour signals signalises a similar mechanism of action and similar clinical effects [13].

Table 5. Effects of VH-04 on motion of the rats.
for the first hour after medicinal product administration. Mean average values are given in ± SEM. Statistical comparison to the results with control were determined using the Wilcoxon, Mann and Whitney U-test.
- Discussion
In this report, we demonstrate that VH-04 is able to alter the EEG-signature of rats, so it could be characterised as a CNS-active medication. We have chosen to go for a holistic method in order to analyse its mode of action systematically in vivo and in vitro. The i.p. injection of VH-04 dose-dependently attenuated delta, theta, alpha 2 and beta 1 waves in the HC, as well as in the FC. In the ST and RF, smaller changes in EEG-signals were observed, but they only reached statistical significance in the ST at 1 ml/kg and in the RF at the highest dosage of 2 ml/kg. The attenuation of alpha 2 waves in the HC and FC, could be interpreted as an activation of the dopaminergic system, because dopamine receptor agonists attenuate alpha 2 waves and dopaminergic receptor antagonists increase alpha 2 waves in this test-system [20]. The prefrontal cortex and the HC formation are densely innervated by dopaminergic afferents, and learning and memory are strongly modulated by acetylcholine and dopamine activity. There is some evidence that the norepinephrine-system affects the incorporation of memories, involving the modulation of synaptic plasticity [32]. Interestingly, both visual and vestibular stimulations have been shown to influence power and frequency of EEG delta, theta and alpha waves in several brain regions of humans [33]. As an outcome of the analysis, the discriminant analysis showed certain similarities in comparison to cognition enhancing cholinergic Metanicotine and Tacrine as evidenced by its positioning in Figure 8, particularly indicated by the close proximity of VH-04 to the reference medicinal product [21]. Many allopathic drugs for vertigo include anticholinergic compounds impairing cognitive functions. However, this is highly speculative and needs to be examined in further studies. Hypothetical effects of VH-04 on cognition are justifiable concerning its effectiveness in the treatment of vestibular vertigo [34], suggesting that information provided by the vestibular system directly affects the HC. Thus, if vestibular input is impeded by bilateral vestibular dysfunction [35] or chronic bilateral vestibular loss [36], spatial memory is impaired in humans. Moreover, vestibular loss results in HC atrophy [36]. If, on the contrary, the vestibular system is stimulated by caloric irrigation or electrical stimulation of the inner ear, cognitive processes are expedited in humans and memory may be enhanced [37] [38]. Another factor is that VH-04 exerts vasorelaxant effects, presumably by regulating smooth muscle cell contraction or relaxation, and there is preliminary evidence for a close relationship between resistance vessel function and neuropsychological performance [39]. Hemispheric malperfusion and altered cerebral blood flow regulation are potentially related to cognitive impairment [40]. For VH-04 subcutaneous microcirculatory improvements caused by VH-04 were demonstrated [5]. Picrotoxin, a GABA antagonist and a secondary metabolite of Anamirta cocculus prevented cognitive impairments in a mouse model of Alzheimer’s disease, according to Yoshiike et al. [41]. Direct exposure of the HC in vitro to several concentrations of VH-04 led to significant increases of the population spike amplitude after SS. Several specific glutamate receptor antagonists were tested to attenuate VH-04 induced increases of the population spike amplitudes. Only compounds interacting with the AMPA and the kainic acid receptor attenuated the effects of VH-04. Dingledine [42] postulates that here is ample evidence that the HC slice preparation in vitro provides an effective method for investigating the communication structure of the brain regarding neurophysiological parameters. Of special interest are surrogate parameters, which reflect cognitive features. Two parameters dominate the analysis of HC function in vitro. The first one consists in the response to the stimulation of the Schaffer collaterals by SS, resulting in the excitation of pyramidal cells. The second one is the initiation of long-term potentiation after theta burst stimulation. Both parameters increase in the presence of memantine, an antidementive medicinal product for the treatment of Alzheimer’s disease, see Dimpfel [43]. VH-04 at rather low concentrations, dilution of 1:4000 of the original preparation, was able to increase these responses. These data corroborate electrophysiological data obtained during the course of in vivo experiments in the second part of the present investigation, where the obtained EPG in rats provides evidence for a stimulatory action of VH-04 on several brain regions. Kainate receptors play a crucial role in the control of synaptic integration and spike transmission efficacy at the HC mossy fiber synapses and are highly implicated in spatial information processing [44]. According to Johnson et al. [45], an AMPA potentiating mechanism might be related also to neuroprotection. In regards to the field potentials and neurotransmission, Dimpfel [15] used more or less specific drugs with known influence on neurotransmission to examine a relationship between field potentials and neurotransmitter activity. Numerous antagonists and agonists of dopamine, norepinephrine, serotonin, glutamate and acetylcholine receptors led to reproducible changes of the frequency content. This indicates that changes in these frequency ranges have a physiological meaning. However, interpretation of these frequency changes is limited, since the activity of several neurotransmitters can change simultaneously in response to a drug. Thus, it is often only possible to assign a dominant change within a particular frequency range to one specific neurotransmitter.
- Conclusion
We conclude that VH-04 has a significant activity on the CNS and that it significantly affects alpha 2 waves in all brain areas. Preliminary observations revealed certain similarities of activity with cognition enhancing drugs, but further experiments are required to support this concept. Preclinical studies are to further investigate the potential effects of VH-04 on memory and cognition, and if these effects can be verified, to further elucidate its mechanism of action.
Authors’ Contributions
- Dimpfel and B. Seilheimer were responsible for the design of the study.
- Dimpfel developed the methodology and was responsible for the interpretation of the results.
- Schombert performed the experiments.
- Seilheimer provided the preparation and pharmacological information of the medicinal product.
- Seilheimer and W. Dimpfel were both involved in writing the manuscript.
Acknowledgements
The Study was financially supported by Biologische Heilmittel Heel GmbH, D-76532 Baden-Baden, Germany. The help of Dr. Stephan Duller, Graz, Austria in preparing the manuscript is highly appreciated.
Conflicts of Interest
The authors declare no conflicts of interest regarding the publication of this paper.
References
|
[1] |
Blakley, B.W. and Goebel, J. (2001) The Meaning of the Word “Vertigo”. Otolaryngology—Head and Neck Surgery, 125, 147-150. |
|
[2] |
Neuhauser, H.K. and Lempert, T. (2009) Vertigo: Epidemiologic Aspects. Seminars in Neurology, 29, 473-481. |
|
[3] |
Fernandez, L., Breinbauer, H.A. and Delano, P.H. (2015) Vertigo and Dizziness in the Elderly. Frontiers in Neurology, 6, 114. |
|
[4] |
Weiser, M., Stroesser, W. and Klein, P. (1998) Homeopathic vs. Conventional Treatment of Vertigo. Archives Otolaryngology—Head and Neck Surgery, 124, 879-885. |
|
[5] |
Klopp, R., Niemer, W. and Weiser, M. (2005) Microcirculatory Effects of a Homeopathic Preparation in Patients with Mild Vertigo: An Intravital Microscopic Study. Microvascular Research, 69, 10-16. |
|
[6] |
Wink, M. (2008) Evolutionary Advantage and Molecular Modes of Action of Multi-Component Mixtures Used in Phytomedicine. Current Drug Metabolism, 9, 996- 1009. |
|
[7] |
Keith, C.T., Borisy, A.A. and Stockwell, B.R. (2005) Multicomponent Therapeutics for Networked Systems. Nature Reviews Drug Discovery, 4, 71-78. |
|
[8] |
Groten, J.P., Butler, W., Feron, V.J., Kozianowski, G., Renwick, A.G. and Walker, R. (2000) An Analysis of the Possibility for Health Implications of Joint Actions and Interactions between Food Additives. Regulatory Toxicology and Pharmacology, 31, 77-91. |
|
[9] |
Wagner, H. and Ulrich-Merzenich, G. (2009) Synergy Research: Approaching a New Generation of Phytopharmaceuticals. Phytomedicine, 16, 97-110. |
|
[10] |
Cassee, F.R., Groten, J.P., Van-Bladeren, P.J. and Feron, V.J. (1998) Toxicological Evaluation and Risk Assessment of Chemical Mixtures. Critical Reviews in Toxicology, 28, 73-101. |
|
[11] |
Berenbaum, M.C. (1989) What Is Synergy? Pharmacological Reviews, 41, 93-141. |
|
[12] |
Chou, T.C. and Talalay, P. (1984) Quantitative Analysis of Dose-Effect Relationships: The Combined Effects of Multiple Drugs or Enzyme Inhibitors. Advances in Enzyme Regulation, 22, 27-55. |
|
[13] |
Dimpfel, W., Schombert, L., Vega-Morales, T. and Wiebe, J. (2016) Neuropharmacological Characterization of Extracts from Rhodiola rosea, Oenothera paradoxa and Paullinia cupana in Comparison to Caffeine. Pharmacology & Pharmacy, 7, 290-303. |
|
[14] |
Dimpfel, W., Dalhoff, B., Hofmann, W. and Schlueter, G. (1994) Electrically Evoked Potentials in the Rat Hippocampus Slice in the Presence of Aminophylline Alone and in Combination with Quinolones. European Neuropsychopharmacology, 4, 151-156. |
|
[15] |
Dimpfel, W. (2006) Different Anticonvulsive Effects of Hesperidin and Its Aglycone Hesperetin on Electrical Activity in the Rat Hippocampus in Vitro. Journal of Pharmacy and Pharmacology, 58, 375-379. |
|
[16] |
Dimpfel, W. (2015) Drug Discovery and Translational Medicine Based on Neuro-Physiological Techniques. A Holistic Approach to Saving Animals. Books on Demand, Norderstedt. |
|
[17] |
Haas, H.L., Schaerer, B. and Vosmansky, M. (1979) A Simple Perfusion Chamber for the Study of Nervous Tissue Slices in Vitro. Journal of Neuroscience Methods, 1, 323-325. |
|
[18] |
Schiff, S.J. and Somjen, G.G. (1985) The Effects of Temperature on Synaptic Transmission in Hippocampal Tissue Slices. Brain Research, 345, 279-284. |
|
[19] |
Lynch, G. and Schubert, P. (1980) The Use of in Vitro Brain Slices for Multidisciplnary Studies of Synaptic Function. Annual Review of Neuroscience, 3, 1-22. |
|
[20] |
Dimpfel, W. (2008) Pharmacological Modulation of Dopaminergic Brain Activity and Its Reflection in Spectral Frequencies of the Rat Electropharmacogram. Neuropsychobiology, 58, 178-186. |
|
[21] |
Dimpfel, W. (2003) Preclinical Data Base of Pharmaco-Specific Rat EEG Fingerprints (Tele-Stereo-EEG). European Journal of Medical Research, 8, 199-207. |
|
[22] |
Dimpfel, W. and Schober, F. (2001) Norepinephrine, EEG Theta Waves and Sedation. Brain Pharmacology, 1, 89-97. |
|
[23] |
Dimpfel, W. (2005) Pharmacological Modulation of Cholinergic Brain Activity and Its Reflection in Special EEG Frequency Ranges from Various Brain Areas in the Freely Moving Rat (Tele-Stereo-EEG). European Neuropsychopharmacology, 15, 673-682. |
|
[24] |
Bennett, D.A., Lehmann, J., Bernard, P.S., Liebman, J.M., Williams, M., Wood, P.L., Boast, C.A. and Hutchison, A.J. (1990) CGS 19755: A Novel Competitive N-methyl-D-aspartate (NMDA) Receptor Antagonist with Anticonvulsant Anxiolytic and Anti-Ischemic Properties. Progress in Clinical and Biological Research, 361, 519-524. |
|
[25] |
Gill, R., Nordholm, L. and Lodge, D. (1992) The Neuroprotective Actions of 2,3-dihydroxy-6-nitro-7sulfamoyl-benzo(F)quinoxaline (NBQX) in Rat Focal Ischaemia Model. Brain Research, 580, 35-43. |
|
[26] |
Szabados, T., Gigler, G., Gacsályi, I., Gyertyan, I. and Lévay, G. (2001) Comparison of Anticonvulsive and Acute Neuroprotective Activity of Three 2,3-benzodiazepine Compounds, GYKI 52466, GYKI 53405, and GYKI 53655. Brain Research Bulletin, 55, 387-391. |
|
[27] |
Pelletier, J.C., Hesson, D.P., Jones, K.A. and Costa, A.M. (1996) Substituted 1, 2-dihydrophtalazines: Potent, Selective, and Noncompetitive Inhibitors of the AMPA Receptor. Journal of Medicinal Chemistry, 39, 343-346. |
|
[28] |
More, J.C., Troop, H.M., Dolman, N.P. and Jane, D.E. (2003) Structural Requirements for Novel Willardiine Derivatives Acting as AMPA and Kainate Receptor Antagonists. British Journal of Pharmacology, 138, 1093-1100. |
|
[29] |
Xi, Z.X., Baker, D.A., Shen, H., Carson, D.S. and Kalivas, P.W. (2002) Group II Metabotropic Glutamate Receptors Modulate Extracellular Glutamate in the Nucleus Accumbens. Journal of Pharmacology and Experimental Therapeutics, 300, 162-171. |
|
[30] |
Kohara, A., Toya, T., Tamura, S., Watabiki, T., Nagakura, Y., Shitaka, Y., Hayashibe, S., Kawabata, S. and Okada, M. (2005) Radioligand Binding Properties and Pharmacological Characterization of 6-amino-N-cyclohexyl-N,3-dimethylthiazolo [3,2-a]benzimidazole-2-carboxamide (YM-298198), a High-Affinity, Selective, and Noncompetitive Antagonist of Metabotropic Glutamate Receptor Type 1. Journal of Pharmacology and Experimental Therapeutics, 315, 163-169. |
|
[31] |
Thomas, N.K., Jane, D.E., Tse, H.W. and Watkins, J.C. (1996) Alpha-Methyl Derivatives of Serine-O-phospate as Novel, Selective Competitive Metabotropic Glutamate Receptor Antagonists. Neuropharmacology, 35, 637-42. |
|
[32] |
Clewett, D., Sakaki, M., Nielsen, S., Petzinger, G. and Mather, M. (2016) Nordrenergic Mechanisms of Arousal’s Bidirectional Effects on Episodic Memory. Neurobiology of Learning and Memory, 137, 1-14. |
|
[33] |
Chen, Y.C., Duann, J.R., Chuang, S.W., Lin, C.L., Ko, L.W., Jung, T.P. and Lin, C.T. (2010) Spatial and Temporal EEG Dynamics of Motion Sickness. Neuroimage, 49, 2862-2870. |
|
[34] |
Morawiec-Baida, A., Lukomski, M. and Latkowski, B. (1993) The Clinical Efficacy of Vertigoheel in the Treatment of Vertigo of Various Etiology. Panminerva Medica, 35, 101-104. |
|
[35] |
Schautzer, F., Hamilton, D., Kalla, R., Strupp, M. and Brandt, T. (2003) Spatial Memory Deficits in Patients with Chronic Bilateral Vestibular Failure. Annals of the New York Academy of Sciences, 1004, 316-324. |
|
[36] |
Brandt, T., Schautzer, F., Hamilton, D.A., Bruning, R., Markowitsch, H.J., Kalla, R., Darlington, C., Smith, P. and Strupp, M. (2005) Vestibular Loss Causes Hippocampal Atrophy and Impaired Spatial Memory in Humans. Brain, 128, 2732-2741. |
|
[37] |
Bachtold, D., Baumann, T., Sandor, P.S., Kritos, M., Regard, M. and Brugger, P. (2001) Spatial- and Verbal-Memory Improvement by Cold-Water Caloric Stimulation in Healthy Subjects. Experimental Brain Research, 136, 128-132. |
|
[38] |
Wilkinson, D., Nicholls, S., Pattenden, C., Kilduff, P. and Milberg, W. (2008) Galvanic Vestibular Stimulation Speeds Visual Memory Recall. Experimental Brain Research, 189, 243-248. |
|
[39] |
Moser, D.J., Hoth, K.F., Robinson, R.G., Paulsen, J.S., Sinkey, C.A., Benjamin, M.L., Schultz, S.K. and Haynes, W.G. (2004) Blood Vessel Function and Cognition in Elderly Patients with Atherosclerosis. Stroke, 35, e369-e372. |
|
[40] |
Marshall, R.S. and Lazar, R.M. (2011) Pumps, Aqueducts, and Drought Management: Vascular Physiology in Vascular Cognitive Impairment. Stroke, 42, 221-226. |
|
[41] |
Yoshiike, Y., Kimura, T., Yamashita, S., Furudate, H., Mizoroki, T., Murayama, M. and Takashima, A. (2008) GABA (A) Receptor-Mediated Acceleration of Aging-Associated Memory Decline in APP/PS1 Mice and Its Pharmacological Treatment by Picrotoxin. PLoS ONE, 3, e3029. |
|
[42] |
Dingledine, R. (1984) Brain Slices. Plenum Press, New York, London. |
|
[43] |
Dimpfel, W. (1995) Effects of Memantine on Synaptic Transmission in the Hippocampus in Vitro. Arzneimittelforschung, 45, 1-5. |
|
[44] |
Sachidhanandam, S., Blanchet, C., Jeantet, Y., Cho, Y.H. and Mulle, C. (2009) Kainate Receptors Act as Conditional Amplifiers of Spike Transmission at Hippocampal Mossy Fiber Synapses. Journal of Neuroscience, 29, 5000-5008. |
|
[45] |
Johnson, K.A., Conn, P.J. and Niswender, C.M. (2009) Glutamate Receptors as Therapeutic Targets for Parkinson’s Disease. CNS & Neurological Disorders—Drug Targets, 8, 475-491. |